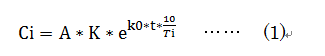
�@
���̐������x
�@
�͂��߂�
���͒Y�_������p�ɂ�萬�����邱�Ƃ��m���Ă���B���̍�p�́A��C���̒Y�_�K�X��t����z�����A��������������A���ɂ��������ɂ����̂ł���ƍl������B���������āA�̐����ɕK�v�ȉh�{�f�́A�Y�_�K�X�̋z�����x�A���̋����A���тɁA���̏Ǝ˖��x�ɂ�荇������Ă���̂ł��낤�B���������h�{�f�̍��������w�����ɂ����̂ƍl����A���ʂȎ��̈���Ă�����������ΐ��̋�������̏Ǝ˂͔�r�I�L�x�ł���̂ł��̉e���͎x�z�I�ł͂Ȃ��ƍl�����A���̔����͒Y�_�K�X�̋������e�����Ă���̂ł͂Ȃ����B�����ŁA��C���̒Y�_�K�X�̋z�������̔����̗��������ɂȂ��Ă���Ƃ̉���̂��ƂɁA��ʓI�ȃK�X�̋z�����x���Q�ƂɎ��̐������x�𐔎������邱�Ƃ����݂��B
�@�����̒萔���قȂ邪�A����́A�������������̂�������ɂ��Ⴂ�ł���A�����̋�����ԁA���z�̏Ǝ˗ʂ̖��A����ɂ́A�C����̈Ⴂ�ɂ����̂ł���̂͗e�Ղɑz�������B
���̐����@
(1) �c�N���@�@�A���A���邢�́A���肵�Ă��琬�����܂�
�@�@�@�@�@�@�@���̊Ԃ́A�Y�_�K�X�̌Œ�͔��ɂ�����肵�Ă���B�@�����������
�@�@�@�@�@�@�@5�`10�N
(2) �������@�@������������i�K�ŁA�Y�_������p�������ɋN����A���̐����������B
�@�@�@�@�@�@�@�}���Ȑ������ŁA���̊ԂɒY�_�K�X�̌Œ肪�N����B
(3) �����݉����@�ߏ�̔ɖɂ��A���̉����ł̒Y�_������p�͑j�Q����A�Y�_�K�X��
�z���̒ቺ���N����B�₪�āA���͐������~�܂�A������ƂȂ�B
(1)�@�@�c�N��
�@�@�@���̊��Ԃ̎��̐������x�́A�����S���ł̃J���}�c�̃f�[�^���Q�l�ɂ����B
�@�@�@�@���c�@�l�@�n�����Z���^�[�@DCM/JI���ƒ������ʃf�[�^�x�[�X
�@�@�@�@�@�����S���X�эĐ��v��x�����ƒ����@�@1999,2000
�@�@�@0�N����Ti�N�܂ł̒Y�_�K�X�̌Œ�ʁ@�ݐ�
�@�@�@�@�@Ti�G���n���J�n�N�@�@�@�@Ti�N�܂ł��c�N��
�@�@
�����ŁA
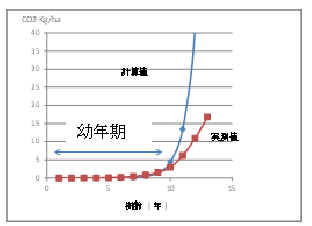
�@�@Ci ; �Y�_�K�X�Œ��(�ݐ�)
A ; ���тɓ��L�̒萔
K ; ���ђn�тɓ��L�̒萔�@
k0
; �c�N���̒Y�_�K�X�Œ葬�x�萔
t ; �o�ߔN��
Ti ;�@�������܂ł̔N��
�@
A 0.00006
K 1
K0 1.12
Ti
10
�@
�@
�@
�@
�@�}-1�@�����S���E�J���}�c�ł̗c�N���̒Y�_�K�X�Œ��(�ݐ�)
(2) ������ �A(3)�@�����݉���
�@�@���̊��Ԃ̐������x�ɂ��ẮA�V��n���X�M�тł̊ϑ��n���Q�l�ɂ����B
�@�@�@�@�X�ыz����������
�@�@�@�@�X�Â���ɂ�����X�ыz�����E�������l���Ȃ�
�@�@�@�@�@�@�@�@�@�@�@�@�]����@�@�@�@����12�N
�@�@�@�@���c�@�l���{�X�їыƐU����@�u�X�щƕK�g�v�ɂ��f�[�^�@�@�@�@�@�@�@�@�@�@�@
�@�@
![]()
�����ŁA
�@
�@Cg
; �������ɂ�����Y�_�K�X�Œ��(�ݐ�)
S; ���т̐����̌��E�@�@�@�@�@�����̑傫���Ɋւ��萔
M; ���ђn�тɓ��L�̒萔�@�@ �����̑����Ɋւ��萔
�@k1:
�������̒Y�_�K�X�Œ葬�x�萔
t ; �o�ߔN��
Tl;�@���̎���
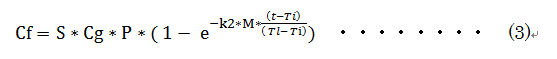
�@�@
�����ŁA
�@
�@Cf
; �o�N�ɂ��Y�_�K�X�Œ�j�Q��(�ݐ�)
S; ���т̐����̌��E
M; ���ђn�тɓ��L�̒萔�@
�@�@�@�@ P ;
���ђn�тɓ��L�̑j�Q�萔
�@�@�@�������j�Q����镔���̊����Ɋւ��萔
k2:
�������̒Y�_�K�X�Œ葬�x�萔
t ; �o�ߔN��
Tl;�@���̎���
���Ȃ݂ɁA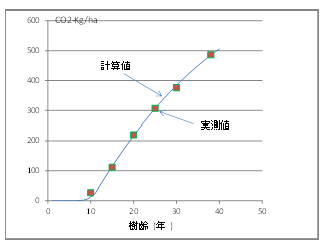
M
6
k1 : 0.2
Tl :
120
�@P
: 8
M
6
k2 :
0.015
�X�т̒Y�_�K�X�̌Œ��(
�ݐ� ) �́A�����ɂ���ċ��߂���B
�@�@�@�@
![]()
�@ (
4 ) �ɏ]���A�M�щJ�т̏ꍇ�̒Y�_�K�X�̌Œ�ʂ𐄒肷��B
�@�@
�@�M�щJ�т̐����̑��x�́A
�c�N���ɂ��ẮA���їт̃����S���̃J���}�c�̔������x�̊��ԂƂ����B�@�@�@�@�@�@
�@�@�@�@�@�@Charlen��̕��Q�l
�@�@�@�@�@�@�@Charlne
Watson, �gForest Carbon Accounting Overview & Principles�h
UNDP 2009
�@�@�@�@���������̒Y�_�K�X�̌Œ�ʂ́A���уX�M�т�1.7�{
�@�@�@�@�@�@�@�@���ؗǖ�@�u�X�̐��Ԋw�v�@�@�@1971�@�@�����o��
�@�@�@�@(4)���ɏ]���A�M�щJ�т̒Y�_�K�X�Œ��(
�ݐ� ) �𐄒肷��B
�@�@�@�@�\-1�@�@�@�@�v�Z�ɗp�����萔
�\�Ɏ������萔��p���Đ��肵���M�їт̒Y�_�K�X�Œ��(
�ݐ� )
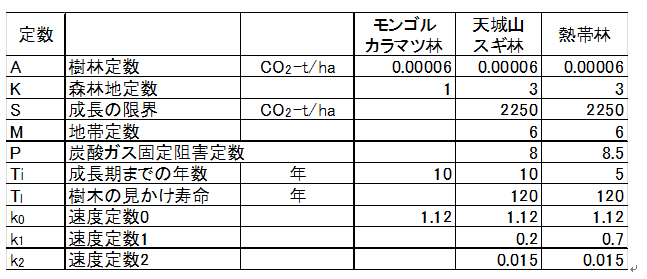
�ȏ�̉�������ɁA�M�щJ�тł̒Y�_�K�X�̌Œ�ʂ����їтł���V��R�̐��т̒Y�_�K�X�Œ�̎����l���琄�����A���̒l�ƁA(�S)�����狁�߂���v�Z�l�̌��ʂ�}-3�@�Ɏ������B
�@�@�@�@
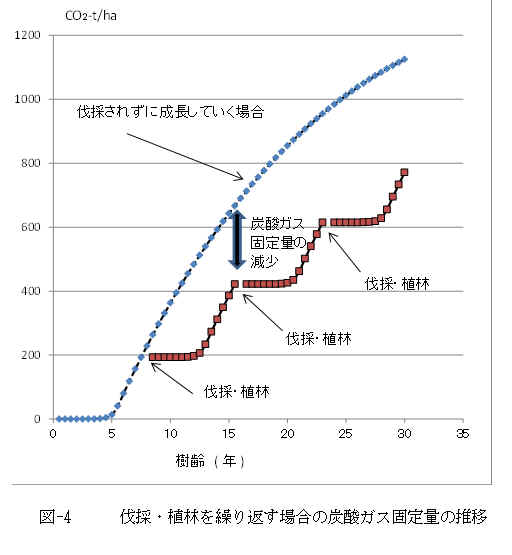
���́E�A�т̉ߒ��ł̒Y�_�K�X�̌Œ�
�@�M�щJ�тŌJ��Ԃ���锰�́E�A�т̉ߒ��ł̒Y�_�K�X�̌Œ�ʂ̐��ڂ́A
�ŏ��̔��̂�7.5�N��Ɏ��s���A�A�т�����Ɠ�x�ڂ̔��̂́A15�`15.5�N��Ɏ��{�����B���̂Ƃ��̒Y�_�K�X�̌Œ�ʂɂ��āA���̂��ꂸ�ɂ��̂܂��т���������ꍇ�ƁA�A�тɂ��A7.5�N��ɍăX�^�[�g���Y�_�K�X���Œ肷��ꍇ�Ƃ��r����ƁA�Y�_�K�X�Œ��(
�ݐ�
)�̍��́A���̎��ƂȂ�B
�@�@�@��Ct
= Ct
(15.5) �|(
Ct
(7.5) + Ct (
8) )
�@�@�@�@�@ = 245 CO2-t/ha
�܂Ƃ�
�@�M�їт̐������x���A���їт̐��т����f���ɂ��āA���̐����̗l�q���V���~���[�V���������B
�@�A���̐����̗l�q�𐔎����������̂ɁARichards�̎����悭�m���Ă���B���̎��́A�A���̐����ɒ��ڂ��āA���̑��x��ϕ����邱�Ƃɂ�萬���̓x���������߂Ă���B�������Ȃ���A���̎��ŕ\������鐬���́A���肩��A�����A�����āA���n�܂ł���̎��ŕ\����Ă���A���肩�琷��ɐ��������i����鐬�����܂ł̊Ԃ̏����i�K�ł́A�����̑��x�͂��傫�߂Ȍ��ʂɂȂ�A�܂��A�������n���ɒB���Đ������~�܂�₪�Ă͖������Ă������ۂȂǂ͗\�����邱�Ƃ��ł��Ȃ��B
�@���̐�����Y�_�K�X������p�̌��ʂƍl����Ȃ�A�����͒Y�_�K�X�̋z���ʂƂ��Ă̖ڈ��ƂȂ邪�A����A�Y�_�K�X�̌Œ�Ƃ����Ӗ��ł́A���̌ċz�ɂ��Y�_�K�X�̔r�o���l������K�v������B���̒Y�_�K�X�̋z���Ɣr�o�̍����Y�_�K�X�̌Œ�ł���̂ŁA�ċz�ɂ��Y�_�K�X�̔r�o�ʂ������Ȃ�A���R�̂��ƂȂ���A�Y�_�K�X�̌Œ�ʂ͌������邱�ƂɂȂ�B�����������Ƃ���A����f���ł́A
�@�@(1)�@�c�N���ł̒Y�_�K�X�̌Œ�ʂ́A�Ǝ��̐��Z��������B
�@�@(2)�@�Y�_�K�X�̌Œ�̐���ɂ́A�Y�_������p�ɂ��z���ƁA�ċz�ɂ��Y�_�K�X��
�r�o���������A���n�ɏ]���Y�_�K�X�̌Œ�ʂ��������鍀�ڂ�����B
���̌��ʁA����f���̏ꍇ�ɂ́A���̐��n��ł́A�Y�_�K�X�̌Œ��(
�ݐ�
)�̌��������Z���邱�Ƃ��ł���B
�������A
�@���݁A���Ƃ��Ă���X�т̔��̂ƐA�т̌J��Ԃ��ɂ��Y�_�K�X�Œ�ʂ̌������́A�������n����ȑO�̒i�K�ł̖��ł���A�\���ɐ����������̂̔��̂Ƃ͑S���قȂ�B�܂�A�Y�_�K�X�̌Œ�ʂɂ��ẮA�����������ԓI�ȊT�O�����邱�Ƃ����ɏd�v�ł��邱�Ƃ����������B
�܂��A�����œ���ꂽ���ꂼ��̒n��ł̎��̐������x��\�������A�A���̐������x�Ɋւ��鐄�Z���Ƃ��Ĉ�ʓI�Ɏ�����Ă���Richards�̎��̕ό`�ł���ƍl���Ă܂��������Ȃ����Ƃ���������
�@
�@
�@